
Бугаев Анатолий Макарович, директор Центра охраны здоровья животных, г.Киев.
Статья помещена на сайте 15 декабря 2013 года.
В течение 17 лет (с 1960 по 1977 год) А.М.Бугаев участвовал в проведении экспериментальных исследований по изучению эндогенных стадий размножения Anaplasma оvis, Toxoplasma gondii и Besnoitia besnoiti на базе лаборатории токсоплазмоза Института зоологии АН КазССР. Исследования проводились совместно с профессором В.Ф. Новинской, СНС В.М. Петешевым и СНС У.Д. Вустиной под руководством академика АН КазССР И.Г. Галузо и профессора Б.П. Всеволодова. Часть результатов, полученных в ходе экспериментальных работ, была опубликована в материалах, представленных на третьем Международном конгрессе протозоологов (1969), а также в книгах «Диагностика токсоплазмоза животных" (1971) и "Жизненный цикл токсоплазм" под редакцией И. Г. Галузо (1974). Часть материалов взяты из докторской диссертации А.М.Бугаева и публикуются впервые.
Светлой памяти моих учителей профессора Бориса Павловича Всеволодова и академика АН.Каз.ССР Иллариона Григорьевича Галузо посвящается этот труд.
БЕЗНОИТИОЗ КРУПНОГО РОГАТОГО СКОТА
Bovine besnoitiosis
(морфология, размножение и систематика Besnoitia besnoiti)
(The relationship between B. besnoiti and several tissues of bovine; the transmission of the disease аnd the life cycle B.besnoiti )
РЕЗЮМЕ
В обзоре дан анализ доступных автору научных работ посвященных изучению жизненного цикла безноитий крупного рогатого скота (Besnoitia besnoiti) и их взаимоотношения с организмом хозяина, начиная с первых публикаций, относящихся к обнаружению паразита в природе в 1912 году и заканчивая оригинальными работами, опубликованными в 2013 году в разных станах мира.
Анализ работ дополняется собственным уникальным архивным материалом, полученным автором на протяжении 40 лет изучения патоморфологии и паразитохозяинных отношений при токсоплазмидозах. В работе большое внимание уделяется паразитохозяинным отношениям и размножению паразитических простейших морфологически и филогенетически сходных с B. besnoiti, которых академик И.Г.Галузо объединил в один отряд Toxoplasmida. Основываясь на достоверном фактическом материале, имеющемся в публикациях и на данных собственных исследований, автор старается показать ошибочность отнесения Besnoitia besnoiti, а равным образом и других токсоплазмид, в одну систематическую группу с эймериями. Во первых, у Besnoitia besnoiti нет половой фазы развития, а для токсплазм она не обязательна; во вторых – у всех токсоплазмид, в бесполой фазе развития и размножения чётко выявляется два этапа размножения: первый этап – это размножение паразитов в лейкоцитах и второй - размножение в клетках РЭС и других клетках организма хозяина; в третьих – для всех токсоплазмид характерным является размножение паразитов как внутриклеточно, так и внеклеточно, а для B. besnoiti и T.gondii размножение внутри ядер клеток организма хозяина. Этих особенностей вполне достаточно, чтобы не объеденять токсоплазмид с эймериями.
Учитывая особенности эндогенного цикла развития токсоплазмид неправомерно применять к безноитиям, токсоплазмам, саркоспоридиям и неоспорам такие термины как «классические и неклассические кокцидии», «типичные и нетипичные коцидии», «цистообразующие кокцидии», «тканевые кокцидии», «облигатно – гетероксенные цистообразующие кокцидии» и т.д. Анализируя данные оригинальных исследований и имеющийся собственный материал, в обзоре особое внимание обращается на то, что паразиты, относящиеся к отряду Toxoplasmida являются, в первую очередь, паразитами форменных элементов крови – сегментоядерных лейкоцитов, лимфоцитов и моноцитов. И здесь речь идёт не о незавершённом фагоцитозе, а о способности токсоплазмид проникать в клетки крови и активно в них размножаться. Кроме того, токсоплазмиды используют клетки крови не только для размножения, но и как транспортные средства для преодоления всех биологических барьеров организма хозяина – кишечный барьер, гематоэнцефалический и даже плацентарный. Показана важная особенность токсоплазмид находить благоприятные условия и активно размножаться в клетках ретикуло – эндотелиальной системы (РЭС) организма хозяина и образовывать в них псевдоцисты. На фактическом собственном материале показана способность токсоплазмид, размножаться как внутриклеточно, так и внеклеточно. Важное место в работе уделено филогенетическому анализу токсоплазмид.
На основании анализа достоверного фактического материала по морфологии паразитических простейших, их жизненным циклам и филогенезу, правомерным в классе Sporozoa создать два подкласса - подкласс Coccidiomorpha с отрядом Coccidiida и семейством Eimeriidae и подкласс Haemosporina с отрядом Toxoplasmida, Haemosporida, Piroplasmida и Adeleida. В отряде Toxoplasmida создать три семейства - семейство Neosporidae с родом Toxoplasmа, Neospora, Besnoitia; семейство Sarcocystidае с родом Sarcocystis и семейство Isosporidae с родом Isospora.
Объединение токсоплазмид с гемоспоридиями поможет ускорить поиск и внедрение в практику более эффективных и безопасных методов лечения не только токсоплазмид, но и других гемоспоридий, таких как малярия, пироплазмоз, тейлириоз и др. заболеваний животных и человека. Практика показала, например, что кокцидиостатики при токсоплазмозе кошек не только бесполезны, но приносят больше вреда, чем пользы.
Есть еще одно обстоятельство, побуждающее исследователей вести поиск новых медикаментов для лечения токсоплазмид: Toxoplasma gondii является близким родственником другого одноклеточного паразита - малярийного плазмодия, говорит профессор Франк Зибер(Frank Seeber) из берлинского Института имени Роберта Коха, Германии. Он считает, что средство, разработанное для подавления токсоплазм, имеет неплохие шансы оказаться эффективным и против плазмодиев.
SUMMARY
This review analyzes the available scientific author of studies on the life cycle beznoity cattle (Besnoitiabesnoiti) and their relationship with the host, since the first publications relating to the detection of the parasite in the wild in 1912 and ending with original research, published in 2012 in different countries around the world.
At the same time used the published and archival materials, obtained by the author for over 40 years of study and pathomorphology and host-parasite relationships with toxoplazmidoz. In this paper attention is paid to host-parasite relationships and reproduction of parasitic protozoa are morphologically and phylogenetically similar to B. besnoiti, which Academician I.G. Galuzo combined in one unit Toxoplasmida. All chapters review the author tries to show the fallacy of attributing Besnoitiabesnoiti, and likewise other toxoplazmid one systematic group with Eimeriidae.
The author considers it wrong to apply to beznoitiyam, Toxoplasma, and undeniable sarkosporidiyam terms such as "classical and nonclassicalcoccidia," "typical and atypical coccidian", "cyst coccidia," "tissue coccidia," "obligate - heteroxenous cyst coccidia", etc. By analyzing data from original research and results of their research, the author draws attention to the fact that parasites are related to the order Toxoplasmida are primarily parasites of blood cells - segmented leukocytes, lymphocytes and monocytes. The blood cells are used by parasites not only to transport them to various apparatus (agencies), but also for its reproduction. The important feature of toksoplazmid is to find favorable conditions and actively multiply in the cytoplasm and cell nuclei reticulo - endothelial system of the host and form in these cells pseudocysts. Reliable fact is the ability of toxoplazmid, multiply both intracellularly and extracellularly. An important place is paid to phylogenetic analysis of toxoplazmid.
Based on an analysis of available publications and its own factual material on the morphology of toksoplazmid, their life cycles and phylogeny, the author considers to be legitimate in the class Sporozoa create two subclasses, - Division Coccidiomorpha with a detachment of the family Eimeriidae and Coccidiida and subclass Haemosporina with detachments Toxoplasmida, Haemosporida, Piroplasmida and Adeleida. In the Detachment Toxoplasmida create three families - the family of the genus Toxoplasma Neosporidae, Neospora, Besnoitia; family Sarcocystidae the genus Sarcocystis and family Isosporidae the genus Isospora.
ОГЛАВЛЕНИЕ
ПРЕДИСЛОВИЕ……………………………………………………………………………………………………………………
ТЕРМИНОЛОГИЯ ИСПОЛЬЗУЕМАЯ В ОБЗОРЕ …………………………………………………………………………….
РАЗДЕЛ ПЕРВЫЙ. ОБЩИЕ СВЕДЕНИЯ О БЕЗНОИТИЯХ И БЕЗНОИТИОЗЕ ЖИВОТНЫХ …
КРАТКАЯ ИСТОРИЧЕСКАЯ СПРАВКА ………………………………………………………………………
МИКРОСКОПИЧЕСКАЯ СТРУКТУРА B.besnoiti ВИДИМАЯ В МАЗКАХ - ОТПЕЧАТКАХ И ГИСТОЛОГИЧЕСКИХ СРЕЗАХ ……………………………………………………………………………
ОБРАЗОВАНИЕ ЦИСТ B.bеsnoiti В ОРГАНИЗМЕ КРУПНОГО РОГАТОГО СКОТА ………
ФАЗА ВНЕКЛЕТОЧНОГО ОБРАЗОВАНИЯ ЦИСТ В. Bеsnoiti …………………………
ФОРМИРОВАНИЕ ОБОЛОЧЕК ЦИСТ ПРИ ВНЕКЛЕТОЧНОМ РАЗМНОЖЕНИЯ И РАЗВИТИИ B.bеsnoiti.
ОСОБЕННОСТИ ФОРМИРОВАНИЯ ЦИСТ B.bеsnoiti ПРИ ВНУТРИКЛЕТОЧНОМ РАЗМНОЖЕНИИ ПАРАЗИТОВ ……………………………………………………
ОБРАЗОВАНИЕ СЕПТИРОВАННЫХ ЦИСТ B.bеsnoiti ……………………………
ГИСТОХИМИЯ ГОМОГЕННОЙ МАССЫ ОБРАЗУЮЩЕЙСЯ ВОКРУГ ЯДЕРНЫХ ПСЕВДОЦИСТ И КОЛОНИЙ B.bеsnoiti ………………
НЕКОТОРЫЕ ВОПРОСЫ РАЗМНОЖЕНИЯ B.besnoiti ………
ГИСТОЛОГИЧЕСКИЕ ИЗМЕНЕНИЯ КОЖИ КРУПНОГО РОГАТОГО СКОТА ПРИ БЕЗНОИТИОЗЕ …………………………………………………………………
ПАТОМОРФОЛОГИЧЕСКИЕ ИЗМЕНЕНИЯ СЕМЕННИКОВ …………
ЗАКЛЮЧЕНИЕ ПО ПЕРВОМУ РАЗДЕЛУ ………………………………
РАЗДЕЛ ВТОРОЙ. ПАРАЗИТИЧЕСКИЕ ПРОСТЕЙШИЕ МОРФОЛОГИЧЕСКИ И ЭВОЛЮЦИОННО СХОДНЫЕ С B.besnoiti …………………………………………
ОБРАЗОВАНИЕ ТКАНЕВЫХ ЦИСТ Toxoplasma gondii В ГОЛОВНОМ МОЗГЕ БЕЛЫХ МЫШЕЙ …………………………
ОБРАЗОВАНИЕ ТКАНЕВЫХ ЦИСТ Toxoplasma gondii В СКЕЛЕТНЫХ МЫШЦАХ БЕЛЫХ МЫШЕЙ ……………………………………………………
ОБРАЗОВАНИЕ ТКАНЕВЫХ ЦИСТ САРКОСПОРИДИЙ В ОРГАНИЗМЕ КРУПНОГО РОГАТОГО СКОТА………………
ВНУТРИКЛЕТОЧНОЕ ОБРАЗОВАНИЕ ТКАНЕВЫХ ЦИСТ Sarcocystis ….
ВНЕКЛЕТОЧНОЕ ОБРАЗОВАНИЕ ТКАНЕВЫХ ЦИСТ Sarcocystis …
ЗАКЛЮЧЕНИЕ ПО РАЗДЕЛУ ……………………
ОБСУЖДЕНИЕ …………………….
О ПОЛОЖЕНИИ B.besnoiti В СИСТЕМЕ ПРОСТЕЙШИХ
ПРЕДИСЛОВИЕ
Безноитиоз крупного рогатого скота (Bovine besnoitiosis) - протозойное, эндемическое заболевание животных тропических и субтропических стран, вызываемое паразитическими простейшими Besnoitia besnoiti. Безноитиоз крупного рогатого скота широко распространён и в Европе, нанося большие экономические потери животноводству (EFSA journal, 2010). В жизненном цикле паразита, до настоящего времени, известен только один способ размножения - бесполый. Заболевание протекает довольно тяжело, характеризуется наклонностью к рецидивированию и преимущественным поражением кожи, склеры, слизистой влагалища у коров и семенников - у быков производителей. Паразиты передаются от животного к животному по горезонтали, путём трансмиссии жалящими насекомыми и по вертикали - половом путём. В 60 – 80 годы прошлого столетия безноитиоз скота был широко распространён в Казахстане и республиках Средней Азии. В настоящее время, в странах бывшего Советского Союза ветеринарные врачи слабо осведомлены о безноитиозе, поскольку заболевание не представляет большого практического значения.
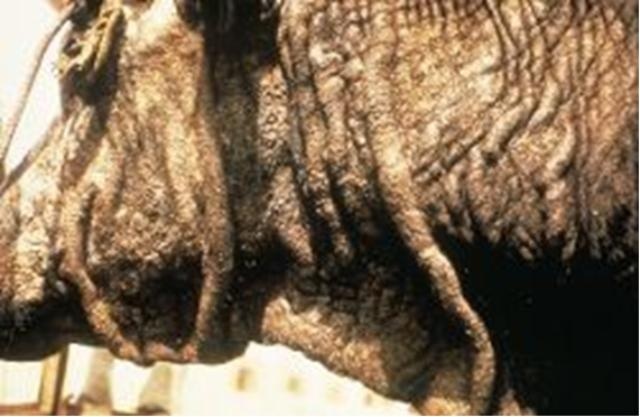
(Фото 1) Типичные макроскопические изменения кожи быка при хроническом безноитиозе. Фотография взята из работы http://medical-dictionary.thefreedictionary.com/besnoitiosis
В жизненном цикле B. besnoiti чётко выделяется две фазы размножения - это фаза строго внутриклеточного размножения, завершающаяся образованием внутрияденых псевдоцист.
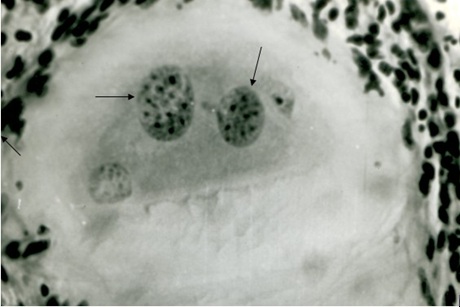
(Фото 2) Гистологический срез кожи 11 месячного бычка при подострой стадии спонтанного безноитиоза. Показаны две ядерных псевдоцисты B.bеsnoiti в гомогенной слизистой массе. Окраска железным гематоксилином и эозином.
и фаза внеклеточного, колониального размножения, завершающаяся образованием тканевых цист размножения (Фото 2 – 3).
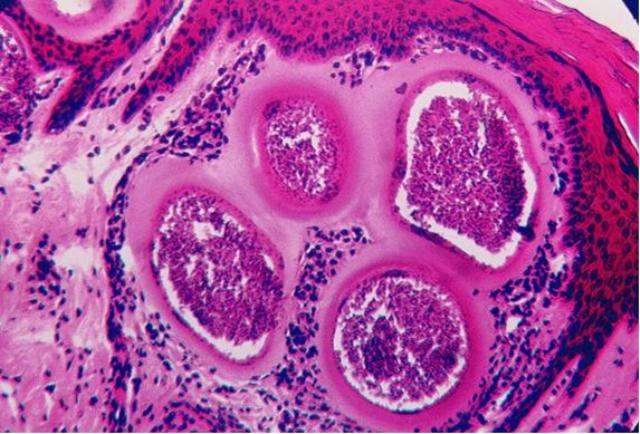
(Фото 3) Гистологический срез кожи коровы 5 лет при хроническом безноитиозе. Под эпидермисом видны хорошо сформированные цисты B.bеsnoiti. Окраска железным гематоксилином и еозином.
Одной из особенностей B.besnoiti является способность их размножаться в белых элементах крови и клетках ретикуло – эндотелиальной системы (РЭС) организма хозяина.
Несмотря на то, что возбудитель заболевания был обнаружен в природе более 100 лет назад, систематическое положение
B. besnoiti до настоящего времени остаётся не ясным. Условно, ещё в 1931 году B.besnoiti были отнесены к группе паразитов неопределённой природы (Якимов, 1931), такими они остаются и до сегодняшнего дня, хотя на сегодняшний день хорошо изучено их эндогенное размножение, морфология и ультратонкая структура. По своей морфологии и электронно – микроскопическому строению B.besnoiti близки к токсоплазмам, саркоспоридиям, эймериям, пироплазмам, малярийному плазмодию и неоспорам.
Однако у B.besnoiti отсутствует половая фаза развития подобная другим токсоплазмидам, гемоспоридиям и кокцидиям. Несмотря на это, безноитии крупного рогатого скота (B.besnoiti) включаются некоторыми исследователями в подсемейство Isosporinae в семействе Eimeridae (Бейер и др.1978), H.Cortes and all.( 2000), а некоторые учёные предлагают уже называть заболевание не безноитиоз, а дерматококцидиоз (Конев, 2011).
Безноитиоз крупного рогатого скота на территории бывшего Советского Союза, впервые был диагностирован в 1959 году профессором Борисом Павловичем Всеволодовым на кафедре патологической анатомии Алма – Атинского ордена Трудового Красного Знамени Зооветиринарного института. И в этом же году результаты исследований были доложены на научной конференции этого же института.

Фото Борис Павлович Всеволодов (1909 – 1981), доктор ветеринарных наук, профессор, заведующий кафедрой патологической анатомии Алма – Атинского зооветеринарного института, заслуженный деятель науки Каз.ССР., многолетний, бессменный вице – президент общества патоморфологов Казахстана.
Борис Павлович Всеволодов – один из ведущих ветеринарных патоморфологов 50 – 70 годов прошлого столетия. После окончания с отличием в 1932 году Московской ветеринарной академии (ныне академия им.К.И. Скрябина) сразу же поступает на работу во Всесоюзный институт гельминтологии, директором которого был основатель гельминтологической школы Советского Союза , академик Константин Иванович Скрябин. В 1937 году, по рекомендации своего учителя Бориса Карловича Боль, Борис Павлович переежжает в Алма- Ату (Казахстан) и взглавляет кафедру патологической анатомии Алма – Атинского зооветеринарного института, которой он заведовал до конца своей жизни. Являясь учеником, одного из основоположников ветеринарной школы патоморфологов Советского Союза Б. К. Боль, Борис Павлович соединил в себе знание и опыт двух школ великих учёных- школы академика Константина Ивановича Скрябина и школы основателя советских патоморфологов Карла Генриховича Боль и всю свою жизнь посвятил изучению патоморфологии гельминтозов сельскохозяйственных, домашних и диких животных. Им был открыт в Казахстане новый для науки гельминт у сайгака, который Борис Павлович назвал его в честь одного из своих учителей – «Скрябинодера». Борис Павлович впервые в СССР, в 1959 году диагностировал неизвестную до того времени болезнь крупного рогатого скота «Безноитиоз» и дал подробное описание патоморфологии этого заболевания. Являясь высоко эрудированным и образованным учёным, кристально чистым и честным к студентам и своим коллегам по работе, Борис Павлович внёс огромный вклад в профессиональное и этическое воспитание многих поколений ветеринарных врачей и учёных Казахстана, и других республик Средней Азии. Под руководством Б.П.Всеволодова защищено около 20 докторских и более 50 кандидатских диссертаций из разных республик бывшего Советского Союза. Он был активным участником многих Республиканских, Всесоюзных и Международных конференций посвящённых патоморфологии болезней сельскохозяйственных животных. Его работы по патоморфологии гельминтозов сельскохозяйственных животных вошли в учебник для высших учебных заведений Советского Союза - «Основы патоморфологии с/х животных», который в советское время переиздавался три раза.
(Автору этого обзора выпала огромная честь быть аспирантом Бориса Павловича, а затем в течение 27 лет работать на кафедре, которую он бессменно возглавлял многие годы).
……………………………………………………………………………………………
Широкомасштабное, целенаправленное и систематическое изучение безноитиоза крупного рогатого скота в СССР начинается с 1961 года, после выступления Б. П. Всеволодова на конференции по вопросам природной очаговости болезней и вопросам паразитологии Казахстана и республик Средней Азии, проходившей 15 -20 сентября в г. Самарканде Узбекской ССР. На этой конференции Б.П.Всеволодов, не только сообщил об обнаружении в Казахстане нового, неизвестного до того времени заболевания крупного рогатого скота - БЕЗНОИТИОЗА, но и подробно продемонстрировал макроскопические и микроскопические изменения в коже и во внутренних органах животных убитых с диагностической целью на мясокомбинате г. Алма-Аты.
Поскольку безноитиоз крупного рогатого скота на территории СССР встречался, в основном, в южных областях Казахстана, то естественно, в изучение этой проблемы активно включились учёные г. Алма – Аты. Все исследования по безноитиозу выполненные в Казахстане, начиная с 1961 года координировались академией наук Каз.ССР. Под общим руководством академика А.Н. Каз. ССР. Иллариона Григорьевича Галузо была разработана республиканская программа, в выполнение которой включились ведущие лаборатории и институты г. Алма – Аты.
На кафедре патологической анатомии зооветеринарного института, под руководством Б.П.Всеволодова, исследования по патоморфологии безноитиоза крупного рогатого скота выполнял доцент кафедры Э.И. Гаппаров; под руководством профессора К.Н. Бучнева, в изучении эпизоотологии безноитиоза крупного рогатого скота и других видов животных, активное участие приняли сотрудники кафедры эпизоотологии - доценты И.У. Увалиев и К.С.Омаров; в Казахском научно – исследовательском ветеринарном институте, в изучение проблемы безноитиоза включились сотрудники лаборатории паразитологии - М. В. Хван (зав.лаборатории), СНС. О.А. Виноградов и Е.Карпухин.
В институте зоологии АН.Каз.ССР., на базе лаборатории токсоплазмоза, которую возглавлял И. Г. Галузо, была организована отдельная группа учёных для изучения биологии и эндогенного развития безноитий при спонтанном и экпериментальном безноитиозе животных. Эту группу возглавил, кандидат биологических наук, СНС лаборатории токсоплазмоза Виталий Михайлович Петешев. В группу В.М. Петешева входил и автор этого обзора. Мне было поручено изучение паразито–хозяинных отношений при спонтанном безноитиозе крупного рогатого скота.

Фото Илларион Григорьевич Галузо (1899 - 1977) — академик АН КазССР.
Илларион Григорьевич Галузо (1899 - 1977) — зоолог, паразитолог, академик АН КазССР, Лауреат Государственной премия СССР 1951. Родился И.Г. Галузо в бывшей деревне Даргейки Сенненского уезда Могилёвской губернии (ныне Витебская область, Сенненский район). В 1926 году окончил Ленинградский ветеринарный институт и был направлен в Среднеазиатский научно исследовательский институт ветеринарии. После трёх месяцев работы в институте Илларион Григорьевич назначается заведующим отделом по паразитарным болезням животных. Работа в Среднеазиатском НИИ и совместные Всесоюзные научно исследовательские экспедиции, возглавляемые видным паразитологом – медиком, академиком МН СССР Е.Н. Павловским, окончательно определили судьбу будущего учёного. В 1933 году Илларион Григорьевич успешно закончил аспирантуру при Институте зоологии АН СССР и после защиты диссертации выполненной под руководством академика Е.Н. Павловского, направляется на работу старшим научным сотрудником Таджикской базы АН СССР.
В 1946 году Илларион Григорьевич, по приглашению академика Каныша Имантаевича Сатпаева переезжает в Алма –Ату и принимает активное участие в организации и формировании биологических институтов АН Каз ССР. После учреждения 1 июня 1946 года Академии наук Казахской ССР, президентом академии был избран К.И. Сатпаев, а Илларион Григорьевич становится директором Института зоологии и первым академиком-секретарём АН. Каз.ССР. В 1956 году Илларион Григорьевич организовывает при Институте зоологии Ан.Каз.ССР., первую в СССР лабораторию токсоплазмоза по изучению биологии и циркуляции токсоплазм в природе. Этой лабораторией Илларион Григорьевич руководил до конца своей жизни.
В Казахстане раскрылся весь талант Иллариона Григорьевича, как учёного и крупного организатора биологической науки. Он является руководителем многих научных экспедиций по Казахстану и в республики Средней Азии. Благодаря его огромному авторитету среди партийной элиты и многолетней, деловой дружбе с академиком АН. СССР Канышем Имантаевичем Сатпаевым И.Г. Галузо удалось провести в Казахстане, Узбекистане и Таджикистане многочисленные научные Всесоюзные и республиканские конференции, симпозиумы и семинары, на которых учёные активно обсуждали научные проблемы по паразитологии, протозоологии и природно – очаговым заболеваниям человека и животных. Проведению таких съездов, конференций и семинаров И.Г. Галузо придавал огромное значение, поскольку считал их важным этапом жизни учёного и хорошей школой для начинающих исследователей. Одной из важных заслуг Иллариона Григорьевича является - создание Казахстанской школы паразитологов, которая объединяла всех учёных, как гельминтологов, так и протозоологов республики.
Илларион Григорьевич Галузо был не только видным учёным, но простым, доступным и скромным Человеком. Он постоянно заботился о научном росте своих учеников и сотрудников. Под его руководством выросло и воспиталось несколько поколений учёных – академиков, докторов, кандидатов наук не только в Казахстане, но и в других республиках бывшего Советского Союза.
За свою активную научную работу и подготовку научных кадров Илларион Григорьевич был награждён Орденами Ленина, Трудового Красного Знамени, медалями и почётными грамотами Верховного Совета КазССР, Золотой медалью имени академика Е. Н. Павловского. За пятитомный труд «Кровососущие клещи Казахстана» Иллариону Григорьевичу в 1951 году была присуждена Сталинская премия. Из всех своих многочисленных наград, Илларион Григорьевич больше всего ценил Золотую медаль имени своего учителя, академика Е. Н. Павловского.
Илларион Григорьевич всю свою жизнь ценил и дорожил деловой и чисто человеческой дружбой с академиком К.И. Сатпаевым. Все ученики Иллариона Григорьевича и учёные Академии Наук Каз.ССР знали об их многолетней дружбе. Поэтому, после смерти И.Г. Галузо было единогласно принято решение, похоронить Иллариона Григорьевича на аллее видных деятелей культуры и науки Казахстана Центрального кладбища г.Алма – Аты, рядом с могилой первого академика – казаха АН.СССР. Каныша Имантаевича Сатпаева.
Основные научные труды Иллариона Григорьевича Галузо
- Кровососущие клещи Казахстана. В 5-ти томах. Алма-Ата, 1946-1953.
- Токсоплазмоз животных. Алма-Ата, 1965.
- Диагностика токсоплазмоза. Алма-Ата, 1971.
- Жизненный цикл токсоплазм. Алма-Ата, 1974.
ПРИМЕЧАНИЕ : Начиная с 1964 и по 1977 год (работая на кафедре патологической анатомии Зооветеринарного института) автору этого обзора посчастливилось выполнять научные исследования в комплексе с замечательными сотрудниками лаборатории токсоплазмоза Института зоологии АН.Каз. ССР. - профессорами Валентиной Фёдоровной Новинской, Виталием Михайловичем Петешевым и СНС Ульяной Дмитриевной Вустиной. Лабораторией в это время, руководил Илларион Григоьевич Галузо и мне судьбой было даровано, на протяжении многих лет, самым тесным образом, общаться с этим видным учёным, простым в обращении и добрым по своему характеру Человеком. Под его руководством мной защищена кандидатская диссертация и написана докторская.
-------------------------------------------------------------------------------------------
В последние 15 – 20 лет безноитиоз крупного рогатого скота в Казахстане и республиках Средней Азии встречается крайне редко и не наносит видимого экономического ущерба хозяйствам. Поэтому, в настоящее время, это заболевание не представляет большого практического интереса для ветеринарных врачей стран СНГ, как это было в 60 – 80 годы прошлого столетия и практически, все исследования по безноитиозу крупного рогатого скота в Казахстане, а равным образом и в странах СНГ, прекращены.
Целью опубликования этого обзора является подведение итогов собственных исследований автором по проблеме токсоплазмид и особенно по вопросу паразитохозяинных отношений при безноитиозе крупного рогатого скота. Автор надеется этой публикацией обратить внимание исследователей к особенностям эндогенного цикла размножения и развития B. besnoiti в организме крупного рогатого скота. На первый взгяд безноитиоз крупного рогатого скота никакого отношения не имеет к токсоплазмозу человека и мелких домашних животных. Однако тщательное знакомство с эндогенным циклом размножения Besnoitia besnoiti проливает свет на многие вопросы размножения токсоплазм и их паразитохозяинные отношения, а равным образом на патогенез токсоплазмоза.
Безноитии крупного рогатого скота занимают особое положение среди других токсоплазмид поскольку к «классическим» кокцидиям, а точнее к эймериям, никакого отношения не имеют. Поскольку такого размножения и взаимоотношения паразита с организмом хозяина нет ни у одного представителя так называемых «истинных кокцидий». Надеюсь, что фактический материал, собранный из различных источников опубликованных в журналах разных стран мира и появившихся в Интернет, а так же данные собственных исследований, поможет учёным изучающим токсоплазмид и вызываемые ими болезни, в решении как теоретических, так и практических вопросов.
ТЕРМИНОЛОГИЯ ИСПОЛЬЗОВАННАЯ В ОБЗОРЕ
1. Токсоплазмиды - это строго ограниченная группа токсоплазмоподобных паразитических простейших (Toxoplasma, Besnoitia, Sarcocystis, Neospora, Frenkelia, Hammondia), являющихся, прежде всего, паразитами лейкоцитов, а затем уже клеток РЭС и других тканей. В фазу генерализации и при хроническом течении болезни, токсоплазмиды способны размножаться как внутриклеточно, так и внеклеточно. Некоторые токсоплазмиды , такие как саркоспоридии и токсоплазмы, могут инвазировать и эритроциты (Фото 1 -8).
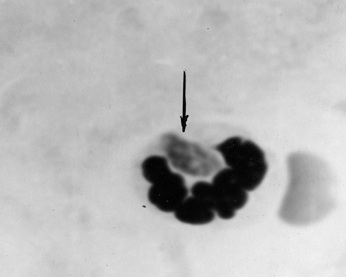
Фото 1. Пластинчатый препарат брыжейки белой мыши через 2 часа после внутрибрюшинного заражения вирулентным штаммом токсоплазм CDN. Показан, хорошо сформированный трофозоит
T. gondii в цитоплазме нейтрофила. Окраска по Романовскому - Гимза.
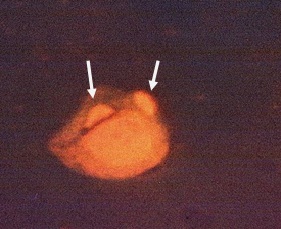
Фото 2. Мазок, приготовленный из брюшного экссудата белой мыши экспериментально заражённой трофозоитами токсоплазм вирулентного штамма CDN. Люминесцентная микроскопия. Препарат обработан акрединовым оранжевым. Показаны два трофозоита в цитоплазме лимфоцита.
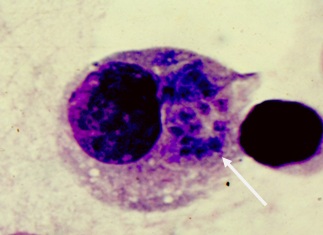
Фото 3. Пластинчатый препарат брыжейки белой мыши на 4 сутки после внутрибрюшинного заражения токсоплазмами вирулентного штамма CDN. Показаны начальные фазы формирования псевдоцисты в цитоплазме моноцита. Окр. По Романовскому – Гимза.
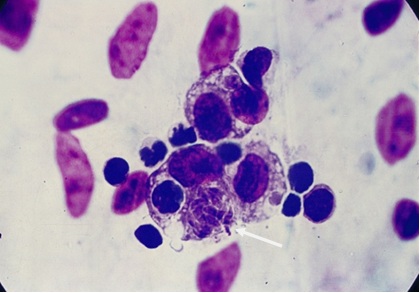
Фото 4. Пластинчатый препарат брыжейки белой мыши. Показаны разные стадии размножения токсоплазм в моноцитах. Окр. по Романовскому - Гимза.
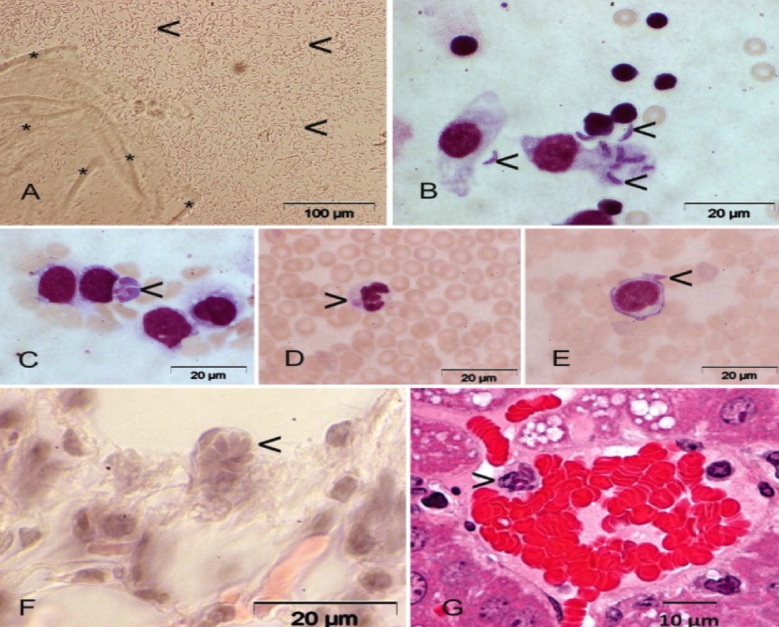
Фото 5 . Показаны трофозоиты безноитий крупного рогатого скота (Besnoitia besnoiti) в мононуклеарах и нейтрофилах перитонеального экссудата белых мышей при экспериментальном их заражении(фиг. В, С,D, Е) , а так же розеткоподобная псевдоциста в гистологическом срезе лёгкого мыши (F) и свободные тахизоиты в очаге кровоизлияния в лёгком.
Fig. 1. Besnoitia besnoiti in a skin biopsy from an infected bull (Animal 70) (A) and in material of an infected γ-interferon knockout mouse (K122) (B–G). (A) Numerous cystozoites (<) are released from tissue cysts (*) in bovine skin after squashing using a mortar and pestle. (B) Peritoneal washing of the B. besnoiti infected γ-interferon knockout mouse with numerous extra- and intracellular tachyzoites (<), Giemsa stained. (C–E) Parasites were observed in blood smear of the GKO mouse (Giemsa stained), either in monocytes (C, <), or in neutrophil granulocytes (D, >) or extracellular (E, <) 5 days post-infection. (F) Parasitophorous vacuole containing B. besnoiti tachyzoites arranged as a rosette (<) in a lung section of the infected GKO mouse, H&E staining. (G) Cluster of tachyzoites (>) in a blood vessel of the liver of the GKO mouse, H&E staining.
Фотографии взяты из работы Германских исследователей « First in vitro isolation of Besnoitia besnoiti from chronically infected cattle in Germany» ☆
- G. Scharesa, , ,
- W. Bassoa, b, c,
- M. Majzoubd,
- H.C.E. Cortese,
- A. Rostaherf,
- J. Selmairg,
- W. Hermannsd,
- F.J. Conrathsa,
- N.S. Gollnickh
- a Friedrich-Loeffler-Institut, Federal Research Institute for Animal Health, Institute of Epidemiology, Wusterhausen, Germany
- b Laboratorio de Inmunoparasitología, Facultad de Ciencias Veterinarias, Universidad Nacional de La Plata, La Plata, Argentina
- c Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Buenos Aires, Argentina
- d Institut of Veterinary Pathology, Ludwig-Maximilians-Universität, Munich, Germany
- e Laboratório de Parasitologica, ICAM, Núcleo da Mitra, Universidade de Évora, Portugal
- f Medizinische Kleintierklinik, Ludwig-Maximilians-Universität, Munich, Germany
- g Inning am Holz, Germany
- h Clinic for Ruminants, Ludwig-Maximilians-Universität, Munich, Germany
http://dx.doi.org/10.1016/j.vetpar.2009.04.033 How to Cite or Link Using DOIPermissions & Reprints
Veterinary Parasitology
Volume 163, Issue 4, 26 August 2009, Pages 315–322
Special Section: EVPC 2008: Veterinary parasitology and climate change.
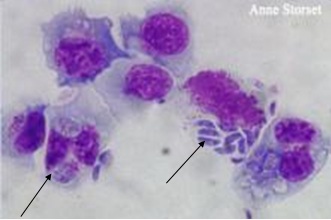
Фото 6. Тахизоиты (трофозоиты) Neospora caninum в лимфоцитах собаки. Фото взято из работы Jackie Barber. Журнал Waltham 1998г. www.sciencedaily.com/.../11021107.
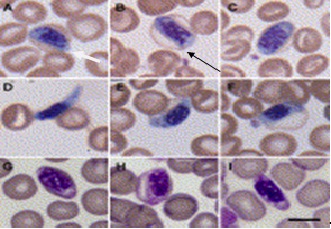
Фото 7. Фото взято из работы Ashlie Hartigana, 1, Bing Y. Zhua, 1, Damien P. Higginsa, Paul J. Canfielda, Jan Šlapetaa, опубликованной в журнале Veterinary Parasitology Volume 159, Issue 2, 5 February 2009, Pages 105–111, выполненая в Австралии. На фотографиях показаны саркоспоридии в эритроцинах.
Fig. 1. Intraerythrocytic parasite (Sarcocystidae; Coccidia; Apicomplexa) in thin blood film. Yellow-bellied glider (Petaurus australis) from Queensland (A–F) and New South Wales (G–I). Intraerythrocytic stages are elongate with an oval shape (A, C and H), some displaying a cylindrical outline (B). Centrally located nucleus is coarsely granular. Emerging parasites take elongated form (I), some with one an extended end (D). Before the emergence of these extracellular forms, the parasite starts to slightly elongate at both ends, with the erythrocyte revealing a piriform shape (E–H). The blood films were stained with Quick Dip (Fronine, Australia)
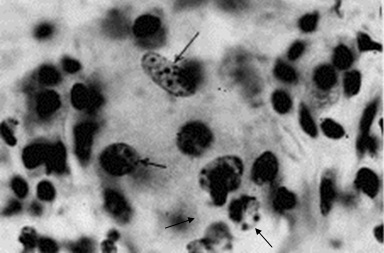
Фото 8. Гистологический срез кожи бычка, подострая фаза безноитиоза. Видны разные стадии размножения безноитий в клетках воспалительного пролиферата - внутриядерное размножение безноитий в гистиоците (показано двумя стрелками) и в цитоплазме лимфоцитов (показано стрелками). В паразитах чётко можно различить ядро и цитоплазму. Окр.Железным гематоксилином и эозином.
Для некоторых токсоплазмид характерным является два независимых цикла рамножения - половой, осуществляемый в организме дефинитивных хозяев и бесполый, осуществляемый в организме промежуточных хозяевах. Благодаря бесполому циклу размножения и развития, токсоплазмиды могут циркулировать среди диких и домашних животных долгое время, практически, бесконечно и таким образом поддерживать существование вида в природе. Для таких токсоплазмид, как Toxoplasma, Neospora, половая фаза развития не обязательна. А у возбудителя безноитиоза крупного рогатого скота (Besnoitia besnoiti) половая фаза развития вообще отсутствует.
Жизненный цикл токсоплазмид, больше сходен с жизненными циклами гемоспоридий и отображает всю эволюцию паразитов этой группы - от свободноживущих жгутиковых до плазмодий маларии человека и безноитий крупного рогатого скота. Поэтому корни происхождения токсоплазмид следует искать, по - видимому, не среди кишечных простейших млекопитающих, а среди жгутиконосцев.
2. Дефинитивный или окончательный хозяин – это организм животного или человека, в котором паразиты размножается половым путём, и достигают половой зрелости. Для токсоплазм это все животные представители семейства кошачьих.
3. Промежуточный хозяин – это организм, в котором паразиты размножаются бесполым путём. Для токсоплазм это все виды млекопитающих и птиц.
4. Транзитный или транспортный хозяин – это организм, в котором паразиты не размножаются, а сохраняются в том виде, в каком они попали к ним. Для токсоплазм это обычно черви, мухи, тараканы, улитки, служащие пищей мелким млекопитающим и птицам.
5. Облигатный хозяин - это хозяин, к которому паразит эволюционно и экологически приспособлен.
5. Факультативный хозяин – это хозяин, к которому паразит филогенетически не приспособлен, обладает в нем слабой жизнеспособностью и низкой плодовитостью. Пример, грызуны для Isospora felis и Isospora rivolta. .
.
6. Моноксенные паразитические простейшие – это паразиты, у которых бесполая и половая фазы размножения проходят в одном и том же организме животного (Пример, эймерии).
7. Поликсенные паразитические простейшие - это паразиты, у которых бесполая и половая фазы размножения проходят в разных видах животных (Пример Toxoplasma gondii и другие токсоплазмиды) .
2. Toxoplasma gondii - Одноклеточный микроскопический паразит, поражающий все виды млекопитающих(включая человека) и птиц. Относится к типу простейших, подтипу Apicomplexa, имеет сложный цикл развития и размножения, как внутри клеток организма хозяина, так и внеклеточно. Именно в жизненном цикле токсоплазм отображена вся эволюция паразитических простейших подкласса Haemosporina. Это подтверждается многими фактами:
Первый - существование и размножение токсоплазм вне клеток – в плазме крови, лимфе, белом веществе головного мозга, между волокнами соединительной ткани и между гладкомышечных волокон.
Второй - размножение токсоплазм внутри лейкоцитов крови - нейтрофилах, лимфоцитах и моноцитах. Имеются сведения о размножении токсоплазм в эритроцитах птиц и эритроцитах куринных эмбрионов при экспериментальном заражении (И.Г.Галузо, С.И.Коновалова, 1974, в книге: Жизненный цикл токсоплазм, ст.84) .
Третий - размножение токсоплазм путём прямого деления материнской клетки на 2 дочерних.
Четвёртый. Наличие жгутиков у микрогамет
Пятый. Копуляция гамет.
Несмотря на такую схожесть токсоплазм с гемоспоридиями и прежде всего с лейшманиями, Toxoplasma gondii имеет свои строгие характеристики, отличающие этих паразитов от других простейших, включая и токсоплазмид.
Первая - Токсоплазмы - это поликсенные паразитические простейшие,
приспособившиеся к паразитированию не только внутриклеточно в клетках крови, гепатоцитах, клетках РЭС и саркоплазме мышечных волокон, но и внеклеточно, в - белом веществе головного мозга, в плазме крови и лимфы, между гладкомышечных и соединительнотканных волокон.
Вторая - В отличие от других паразитических простейших, у токсоплазм, как в организме дефинитивных, так и промежуточных хозяев, нет чётко детерминированных стадий бесполого размножения. Бесполое размножение может происходить бесконечно долго, циркулируя между животными, минуя половую фазу.
Пимечание. У всех спорообразующих паразитов, даже гемоспоридий, бесполое размножение не проделывается бесконечное число раз, у токсоплазм оно может продолжаться бесконечно. Доказательство - поддержание десятилетиями в лабораторных условиях вегетативных форм токсоплазм на чувствительных лабораторных животных и в культуре клеток. Для поддержания вида в природе споровикам обычно необходима следующая за бесполым размножением стадия полового развития. Для токсоплазм она не обязательна.
При размножении токсоплазм даже в кишечнике представителей семейства кошачьих, особи токсоплазм одного и того же типа (по терминологии Dubey и Frenkel, 1972) могут воспроизводить себе подобных, тогда как у кокцидий такая способность отсутствует. У кокцидий, шизонты, например, первой генерации не могут воспроизвести себе подобных, они должны дать потомков (мерозоитов), относящихся уже к другой генерации.
Третья – У всех споровиков начальной стадией цикла развития и размножения является спорозоит, у токсоплазм чаще – трофозоит, мерозоит, брадизоит и только иногда спорозоит.
Четвёртая. Отсутсвие прямого цикла размножения в кишечнике дифинитивного хозяина - от изосоры (ооцисты) до изосоры (ооцисты).
Пятая. Способность любой фазы развития и размножения (кроме гаметоцитов и неспорулированных зигоцист) заражать животных и человека при любом пути проникновения в организм, даже интрацеребрально.
Шестая. Способность изоспор токсоплазм заражать организм животных и человека как перорально, так и парентерально - подкожно, внутрибрюшинно, внутримышечно, интрацеребрально, чего не бывает у так называемых «истинных кокцидий».
Седьмая. Дуализм развития и размножения брадизоитов токсоплазм в организме дефинитивных хозяев - давть начало половой фазе размножения в эпителии кишечника и одновременно внекишечной, бесполой (фазе трофозоита и цист размножения) в других органах и тканях того же животного.
Восьмая. Отсутствие у дефинитивных хозяев половой фазы размножения при пероральном введении трофозоитов вирулентных штаммов, например, штамма RH. В то время как общее заражение животных происходит.
Девятая. Способность развиваться и размножаться в организме многих видов животных, а также человека.
Десятая. Тройной механизм передачи от донора реципиенту - первый, путём контаминации, через цисты, трофозоиты и изоспоры, которые проникают в организм животных как перорально, так и парэнтерально; второй, внутриутробно - от матери к плоду через плаценту; третий, путём хищничества - хищник - жертва.
Одинацатая. Животные заражаются не только от особей того же вида, но и от разных видов млекопитающих и птиц.
2.Токсоплазмидозы - в широком смысле - это заболевания вызываемые токсоплазмидами. По нашей систематике - паразиты, относящиеся к отряду Toxoplasmida. Однако в практической деятельности, диагноз токсоплазмидоз никто из врачей не пишет. И это правильно. Хотя, теоретически, такой термин имеет право на существование, как, например, термин «кокцидиозы» объединяющий все заболевания вызываемые паразитами, относящимися к отряду Coccidiida. В основу названия токсоплазмид (если диагноз пишется на русском языке) берётся родовое название паразита и добавляется окончание (оз). Например, в основу названия болезни вызываемой токсоплазмами, берётся родовое название паразита -Toxoplasma и добавляется (оз), в результате получается название болезни - токсоплазмоз. Так же поступают и с другими болезнями вызываемых токсоплазмидами - безноитиоз, неоспороз, гаммондиоз, френкелиоз, изоспороз, саркоцистоз
Если хотят, например, при обнаружении в кале спороцист саркоспоридий подчеркнуть происхождение спороцист, то пишут Sarcocystis bovifelis или Sarcocystis bovicanis и т.д. Но такая терминология приемлема при экспериментальных исследованиях, когда точно известно, от какого животного берутся тканевые цисты. На практике же, при обнаружении спороцист в кале собаки или кошки, врачу трудно определить, какому виду саркоспоридий принадлежат спороцисты, к бычьим, овечьим или другим видам сельскохозяйственных и диких животных. Да и вообще для лечения животных (собак и кошек) это определение большого значения не имеет. Поэтому, правильнее будет, если врач при обнаружении спороцист в кале животного, в основу написания диагноза, возьмёт родовое название возбудителя - Sarcocystis и напишет в заключении - САРКОЦИСТОЗ.
3. Эймерии или «типичные кокцидии» - это большая группа строго внутриклеточных паразитических простейших животных и человека. Эймерии паразитируют только в эпителиальных клетках кишечника, жёлчных протоков и эпителии почек. В отличие от токсоплазмид, у эймерий строго детерминированы все стадии бесполого и полового развития. Они размножаются только путём шизогонии, с последующим распадом шизонта на мерозоиты. У эймерий, после нескольких бесполых поколений обязательно наступает половой процесс. (Токсоплазмы же, могут бесконечно долго размножаться бесполым путём, как в организме промежуточных, так и дефинитивных хозяев и поддерживать популяцию в природе, минуя половую фазу развития). У эймерий инвазионными являются только спорулированные ооцисты, содержащие по четыре спороцисты, в каждой, из которых формируется по два спорозоита. Заражение может произойти только при пероральном попадании спорулированных ооцист в организм животного или человека. Эймерии - это моноксенные паразитические простейшие.
4. Кокцидиозы или эймиреозы - это заболевания вызывемые эймериями или «типичными» кокцидиями - паразитами, относящимися к отряду Coccidiida.
Примечание. В предлагаемой нами систематике токсоплазмид, паразитические простейшие, формирующие в зигоцистах по две споры с четырьмя спорозоитами, выделены в отдельное семейство отряда Toxoplasmida. Это было продиктовано тем, что у таких изоспор как
I. felis и I. rivolta, в организме кошек и собак были обнаружены (Dubey, Frenkel, 1972), наряду с кишечными стадиями бесполого размножения, внекишечные стадии развития паразитов(Развития, но не размножения!). Этот цикл вегетативного развития сходен (но не идентичен!) с таковым у токсоплазм и неоспор.
5. Трофозоит или тахизоит - быстро размножающаяся стадия токсоплазмид. Тахизоиты могут размножаться как внутриклеточно, так и внеклеточно, как в цитоплазме клеток, так и в их ядрах. В стадию трофозоита обеспечивается быстрое увеличение числа паразитов в организме хозяина. Питание трофозоитов происходит за счёт гидролизированной структуры цитоплазмы или ядра инвазированной клетки, или окружающей трофозоит ткани организма хозяина, если паразит находится внеклеточно.
6. Гомогенизированная масса - это клетки и ткани организма хозяина, подвергшиеся гидролизу ферментами выделяемыми токсоплазмидами. В конечном счёте, гомогенизированная масса представляет собой низкомолекулярные блоки, используемые паразитами для дальнейшего синтеза собственных нуклеиновых кислот, белков, углеводов и других веществ, необходимых как для питания, паразитов, так и формирования цистной оболочки. Гидролизируя окружающею ткань, токсоплазмиды обеспечивают себя необходимым питательным материалом для развития и размножения (Фото 9 - 12).
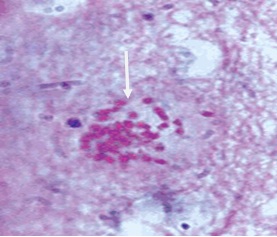
Фото 9. Гистологический срез коры головного мозга белой мыши экспериментально зараженной токсоплазмами слабовирулентного штамма "LЕI". Показана гомогенизированная ткань (гидролиз) белого вещества мозга вокруг внеклеточных брадизоитов. Ок. по Мак Манусу.
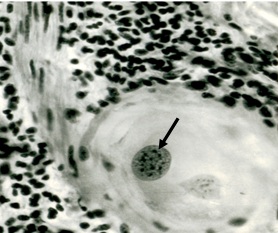
Фото 10. Гистологический срез кожи бычка спонтанно зараженого токсоплазмоподобным паразитическим простейшим Besnoitia besnoiti.Показано ядро - псевдоциста гладкомышечной клетки, в котором наблюдается размножение паразитов. Вокруг псевдоцисты видна широкая зона гидролизированной окружающей ткани - гладкомышечных волокон и ядер воспалительного инфильтрата. Окраска железным гематоксилином и эозином.
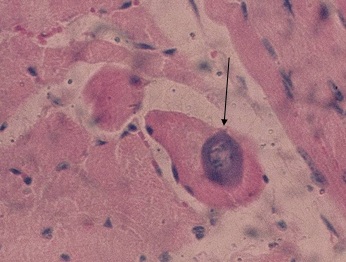
Фото 11. Гистологический срез миокарда правого желудочка зебу. Куба, 1981 год. Естественное заражение животного саркоспоридиозом. В саркоплазме мышечного волокна виден делящийся метроцит, вокруг которого просматривается зона гомогенизированной саркрплазмы окрашенной базофильно. Окр. Гематоксилин – эозином.
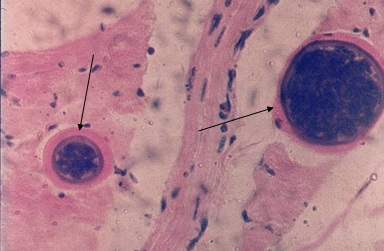
Фото 12. Гистологический срез миокарда зебу. На фотографии показаны две формирующиеся цисты в саркоплазме мышечных волокон Sarcocystis sp. Слева на фотографии видна небольшая колония паразитов с широкой оболочкой и двумя мембранами – внутренняя отделяет делящихся паразитов от широкого базофильного слоя и наружная отделяет широкий базофильный слой от изменённой (гидролизированной) саркоплазмы мышечного волокна. Гидролизированная саркоплазма вокруг колонии паразитов окрашена интенсивно эозином. Справа, показана колония паразитов, в которой видно большое количество делящихся паразитов окрашивающихся базофильно. Вокруг колонии, базофильный слой оболочки цисты сохранен, а гомогенизированная саркоплазма мышечного волокна сдавлена и представляет собой небольшой эозинофильный ободок. Окр. железным гематоксилином и эозином.
Отличием гомогенизированной саркоплазмы мышечного волокна расположенной вокруг метроцитов Sarcocystis, от гомогенизированной окружающей ткани образующейся вокруг ядерных псевдоцист Besnoitia besnoiti заключается в том, что гомогенизированная саркоплазма даёт резко положительную реакцию по Браше на РНК. В то время как гомогенизированная масса вокруг безноитиозных псевдоцист, хотя и содержит РНК, но всё же в ней преобладают нейтральные мукополисахариды (Фото 13 ). Следует особо отметить, что брадизоиты, например, токсоплазм и безноитий содержат больше амилопектина, чем РНК, в то время, как в цитоплазме брадизоитов саркоспоридий обнаруживаются только следы амилопектина. Тогда как РНК заполняет, практически всю цитоплазму паразитов.
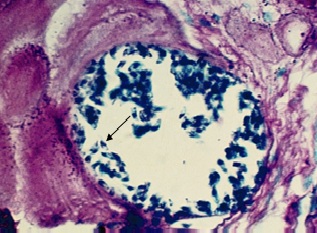
Фото 13. Гистологический срез скелетной мышцы зебу. Показан поперечный срез тонкостенной цисты саркоспоридий. Реакция цитоплазмы паразитов на амилопектин негативная, тогда как саркоплазма мышечных волокон даёт положительную реакцию на нейтральные мукополисахариды. Окр. По Мак - Манусу.
- 7. Паразитофорная вакуоль - это зона гидролиза цитоплазмы инвазированной клетки или ткани окружающей паразита, где концентрируются низкомолекулярные блоки для дальнейшего синтеза питательных веществ необходимых для паразита (Фото 14- 15).
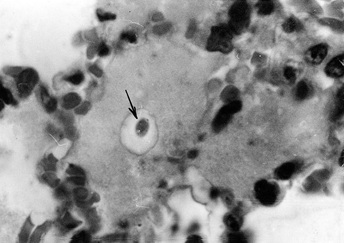
Фото 14. Гистологический срез лёгкого щенка, павшого на 15 сутки от острого экспериментального токсоплазмоза. В серозном экссудате видна токсоплазма (тахизоит) с хорошо выраженной паразитофорной вакуолью. Окр. Гематоксилин - эозином.
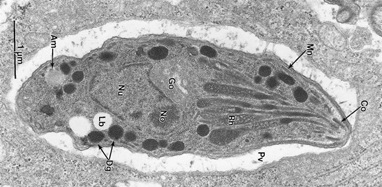
Фото 15. На электронномикроскопической фотографии показана паразитофорная вакуоль вокруг тахизоита T. gondii расположенной в цитоплазме клетки перитонеального экссудата (какой клетки авторы не указывают). Фото взято из работы J. P. Dubey,1,* D. S. Lindsay,2 and C. A. Speer3« Structures of Toxoplasma gondii Tachyzoites, Bradyzoites, and Sporozoites and Biology and Development of Tissue Cysts»† Работа появилась в Интернете в 2003 году.
Transmission electron micrograph of a tachyzoite of the VEG strain of T. gondii in a mouse peritoneal exudate cell. Am, amylopectin granule; Co, conoid; Dg, electron-dense granule; Go, Golgi complex; Mn, microneme; No, nucleolus, Nu, nucleus; Pv, parasitophorous (more ...)
8. Колониальная паразитофорная вакуоль - это внеклеточная зона гидролизированной ткани организма хозяина, образовавшаяся путём гидролиза вокруг колонии паразитов и служащая питательным субстратом для внеклеточных, свободных брадизоитов, а так же паразитов расположенных внутри ядер и цист (Фото 16 ).
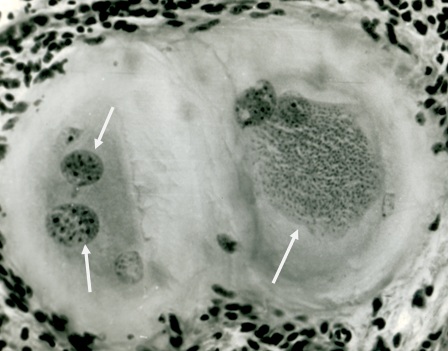
Фото 16. Гистологический срез кожи 11 месячного бычка при подострой стадии спонтанного безноитиоза. Показаны две одновременно формирующиеся тканевые цисты B.bеsnoiti в гидролизированной окружающей ткани. В одной из них, слева на фотографии, видно три ядерных псевдоцисты. В другой – три ядерных псевдоцисты и большая колония брадизоитов в гомогенной массе. Вокруг колонии видна уже мембрана, которая, с одной стороны, удерживает колонию компактно и граничит с питательной массой – формирующейся колониальной паразитофорной вакуолью. Окраска железным гематоксилином и эозином.
У цист с толстыми оболочками, колониальная паразитофорная вакуоль формируется в среднем слое оболочки. И, по – видимому, пока этот слой существует циста не дегенерирует. После истончения этого слоя в цисте наблюдается дистрофия цистозоитов и такие цисты подвергаются атаке макро, - и микрофагов и они погибают, а на их месте образуются гранулёмы (Фото 17 – 21)
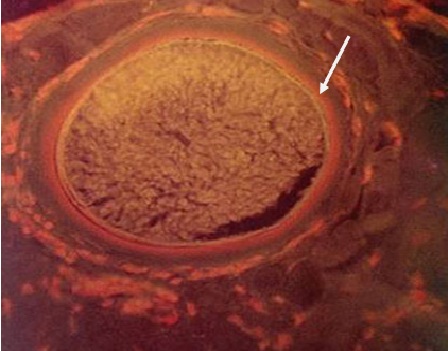
Фото 17. Гистологический срез кожи бычка, подострое течение безноитиоза. Препарат обработан акридиновым оранжевым. Внутренний слой (колониальная паразитофорная вакуоль) флюоресцируют оранжевым светом, что свидетельствует о содержании в ней большого количества РНК.
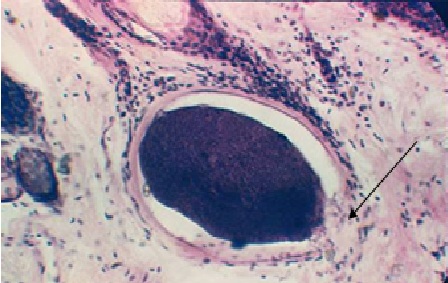
Фото 18. Гистологический срез дермы коровы больной хроническим безноитиозом. Показана дистрофия цистозоитов, деструкция оболочки цисты и проникновение клеток воспалительного инфильтрата внутрь цисты. На фотографии хорошо видна внутренняя мембрана оболочки цисты, которая удерживает колонию погибающих паразитов. Колониальная паразитофорная вакуоль представлена в виде тонкого базофильного ободка. Окр. Гематоксилин – эозином.
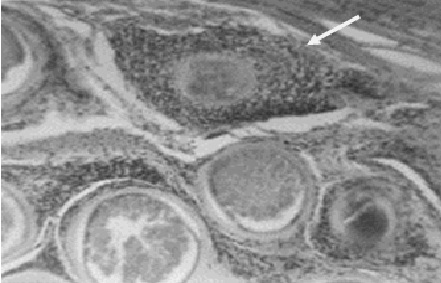
Фото 19. Гистологический срез дермы коровы при хроническом течении безноитиоза. Наряду с хорошо сформированными цистами видна погибающая циста, окружённая большим количеством клеток организма хозяина. Окр. Гематоксилин – эозином.
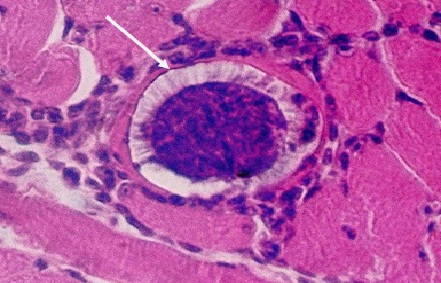
Фото 20. Гистологический срез скелетной мышцы буйволицы породы зебу погибшей от «Внезапной смерти» в провинции Баямо Республики Куба. Показана дистрофия толстой оболочки цисты саркоспоридий при естественном заражении и проникновение в полость цисты микрофагов. Окр. Гематоксилин - эозином.
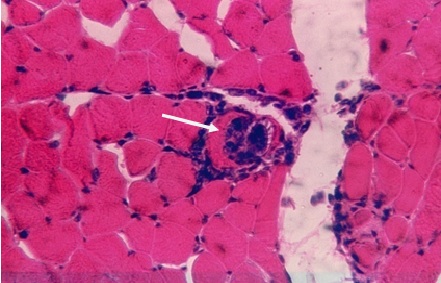
Фото 21. Гистологический срез скелетной мышцы буйволицы породы зебу погибшей от «Внезапной смерти». Показана гибель толстостенной цисты саркоспоридий и замещение её клетками пролиферата. Окр. Гематоксилин - эозином.
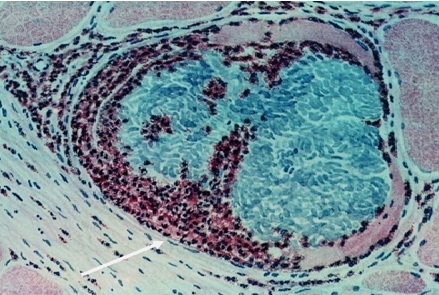
Фото 22. Фото взято из учебника по патологической анатомии сельскохозяйственных животных Ниберле и Корса, 1980. На рисунке показана гибель цистозоитов саркоспоридий и проникновение макро, - и микрофагов внутрь цисты.
У токсоплазм и токостенных цист саркоспоридий гибель цист, по - видемому, связана с другими факторами нарушающими питание многочисленной колонии токсоплазм внутри цисты, а именно с утончением и исчезновением прецистного гомогенного слоя (колониальной паразитофорной вакуоли) (23-25).
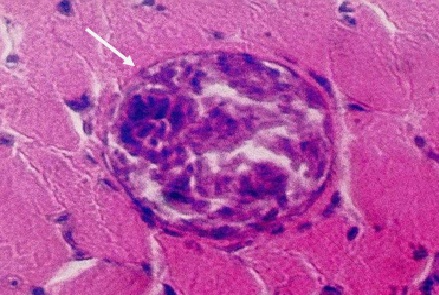
Фото 23. Гистологический срез скелетной мышцы буйволицы породы зебу погибшей от «Внезапной смерти» в провинции Баямо Республики Куба. На фотографии показана тонкостенная циста саркоспоридий. Местами хорошо видно утончение прецистного гомогенного слоя (колониальной паразитофорной вакуоли) и проникновение, в местах утончения прецистного слоя, фагоцитов внутрь цисты. Окр. Гематоксилин - эозином.
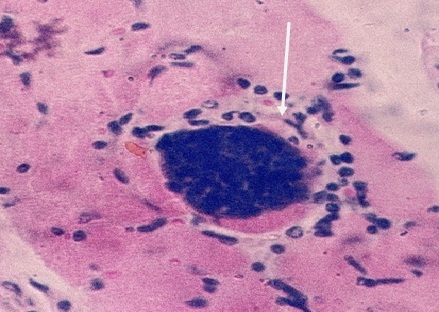
Фото 24. Гистологический срез скелетной мышцы бычка зебу погибшего от «Внезапной смерти» в провинции Баямо Республике Куба. Показана погибающая тонкостенная циста саркоспоридий и проникновение внутрь цисты макро, и микрофагов.
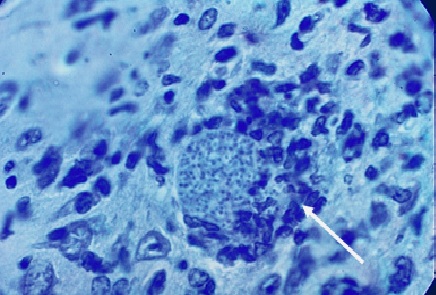
Фото 25. Гистологическаий срез головного мозга белой мыши экспериментально заражённой слабовирулентным штаммом токсоплазм «LEI». Показан момент гибели и фагоцитоза цисты. Проникновение микро, - и макрофагов наблюдается в участке исчезновения прецистного питательного слоя. Окраска железным гематоксилином.
9. Колония токсоплазмид - это группа внеклеточно размножающихся и развивающихся брадизоитов формирующих цисту (Фото 26).
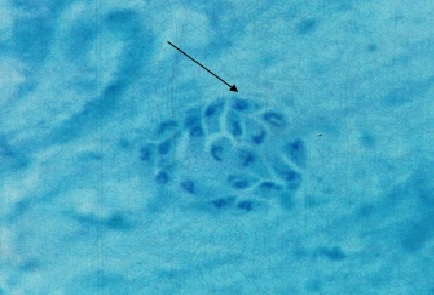
Фото 26. Мазок - отпечаток головного мозга белой мыши заражённой слабовирулентным штаммом токсоплазм «LEI» и убитой через 10 суток после заражения. Показана небольшая, внеклеточная колония токсоплазм в белом веществе стволовой части мозга. Вокруг колонии паразитов оболочка ещё отсутствует. Окраска железным гематоксилином с подкраской метеленовым синим.
Тканевая циста токсоплазмид - это сферическая или продолговато – овальная, внеклеточная колония, медленно размножающихся бесполым путём паразитов, окружённая чётко контурированной оболочкой. Тканевая циста это одна из фаз размножения и развития паразитов. Тканевые цисты, например, у токсоплазм образуются не только в организме животных и человека, но и в культуре клеток, где нет никаких антител(Gustavson at all., 1954; Hogan at all.,1960, 1961; Beverley,1969; Галузо, Коновалова, 1974; Baron S.1996).. Поэтому говорить, что тканевая циста образуется как результат защитных механизмов организма хозяина нет оснований. Циста - это «кокан» или «дом» построенный самими паразитами за счёт материала организма хозяина. Грануляционная ткань организма хозяина при этом никакого участия не принимает (Фото 27 -34).
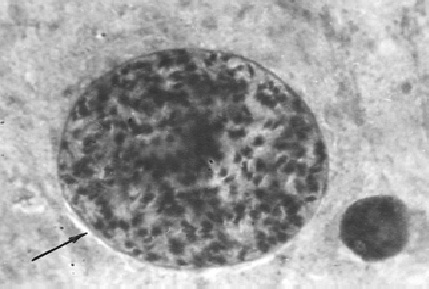
Фото 27. Мазок - отпечаток головного мозга белой мыши. Показана циста токсоплазм с хорошо контурированной оболочкой. Внутри цисты видно множество брадизоитов. Никакой грануляционной ткани вокруг цисты не видно. Окраска железным гематоксилином и эозином.
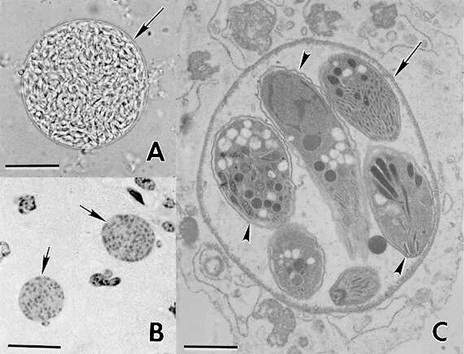
Фото 28. A.Tissue cyst freed from mouse brain. Note a thin (arrow) cyst wall enclosing hundreds of bradyzoites. Unstrained. Bar = 20 μm.
B.Two tissue cysts (arrows) in section of brain. Hematoxylin and eosin stain. Bar = 20 μm.
C.Transmission electron micrograph of a small tissue cyst in cell culture. Note thin cyst wall (arrow) enclosing 6 bradyzoites (arrowheads). Bar = 1.0 μm. (Courtesy of Dr. D.S. Lindsay, Auburn University, Auburn, AL.)
На фотографиях показаны внеклеточные цисты токсоплазм в головном мозге экспериментально заражённой белой мыши и внутриклеточное формирование цисты в культуре клеток.

Фото взято из статьи Baron s.(1996).
Medical Microbiology. 4th edition.
Baron S, editor.
Galveston (TX): University of Texas Medical Branch at Galveston 1996.
Copyright © 1996, The University of Texas Medical Branch at Galveston.NCBI Bookshelf. A service of the National Library of Medicine, National Institutes of Health.
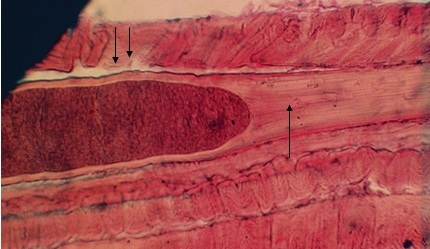
Фото 29. Продольный гистологический срез скелетной мышцы зебу. Показана большая колония паразитов (циста) саркоспоридий внутри мышечного волокна, с хорошо сформированной тонкой цистной оболочкой. Саркоплазма инвазированного волокна превращена в гомогенную массу (Показано стрелкой). Сарколема при этом хорошо сохранена, базофильно окрашена и никакого участия в образовании цистной оболочки не принимает (Показано двумя стрелками). Окраска гематоксилин - эозином.
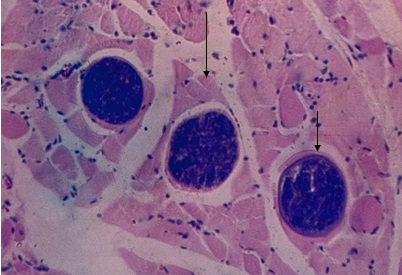
Фото 30. Гистологический срез миокарда правого желудочка зебу. На фотографии показаны три хорошо сформированные цисты саркоспоридий. В цисте расположенной внизу справа (показано стрелкой), базофильный слой ещё хорошо просматривается и видно, что сарколемма мышечного волокна никакого участия в формировании оболочки цист не принимает. В средней и верхней цисте базофильный слой истончён, саркоплазма мышечного волокна гомогенизирована и прижата к сарколеме мышечного волокна. Местами создаётся впечатление, что у этих цист наружным слоем является сарколема мышечного волокна. Окр. Железным гематоксилином и эозином.
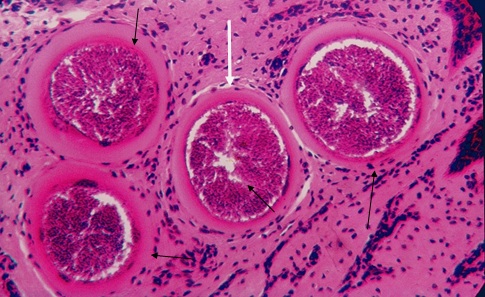
Фото 31. Гистологический срез дермы коровы при хроническом безноитиозе. Показаны хорошо сформированные внеклеточные цисты B.bеsnoiti с множеством брадизоитов. Вокруг цист видна только гомогенная эозинофильная масса. Соеденительная ткань никакого участия в этом не принимает, кроме как ткань подвергшаяся гидролизу. Окр. Гематоксилин – эозином.
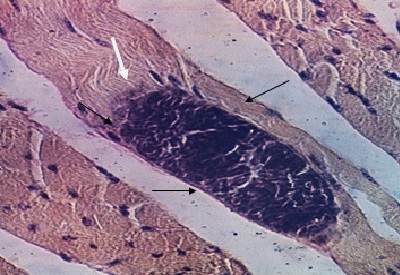
Фото 32. Гистологический срез скелетной мышцы 14 месячного бычка породы зебу павшего от «Внезапной смерти». Показан продольный срез атрофированного пучка мышцы с формирующейся в нём внеклеточной цисты саркоспоридий. Окр. Железным гематоксилином и эозином.
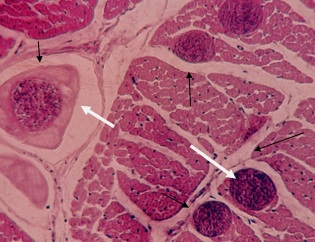
Фото 33. Гистологический срез миокарда правого желудочка зебу. Показаны хорошо сформированные цисты саркоспоридий в мышечных волокнах (Показано стрелками) и в нервном волокне (Показано двумя стрелками). Окр. Гематоксилин - эозином.
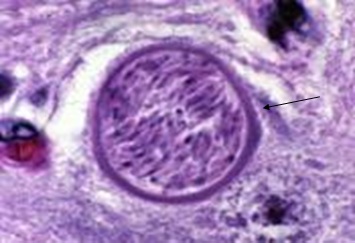
Фото 34. Внеклеточная, толстостенная циста неоспор в головном мозге двухгодичного роттвелера. Фото взято из работы Jackie Barber. Журнал Waltham 1998г.
www.sciencedaily.com/.../11021107.
8. Брадизоиты - это медленно, внеклеточно размножающие паразиты, формирующие тканевую цисту. По морфологии они бывают двух типов. Одни с центральным расположением ядер, другие с ядрами расположенными у тупого края, последние напоминают вид парашутика. Теоретически и практически, брадизоитами можно называть и паразитов находящихся внутри цист, поскольку паразиты продолжают медленно размножаться и после формирования оболочки вокруг колонии паразитов. Однако удобнее и понятнее, паразитов в сформировавшейся цисте называть цистозоитами, как это принято большинством исследователей.
Таким образом, при светооптическом исследовании давно было замечено, что тканевые цисты токсоплазм содержат паразитов находящихся на разных стадиях развития(Фото 35). Как и у других токсоплазмид, в цистах происходит подготовка паразитов к половому развитию и размножению. Поэтому от зрелости цист зависит препатентный и патентный период в кишечнике кошек.
Исследователи, изучавшие цисты токсоплазм и их взаимоотношения с организмом хозяина, никогда не видели, что брадизоиты вызывают некроз окружающей ткани. Брадизоиты могут вызвать только гомогенезацию или вернее гидролиз окружающих клеток и тканей, но не некроз. Гидролизуя окружающею ткань брадизоиты тем самым подготавливают окружающею среду, для последующего синтеза из неё питательных веществ необходимых паразитам находящихся в колонии. Брадизоиты отличаются от тахизоитов не только антигенной структурой и темпами размножения, но и метаболизмом, тропностью и патогенностью. Например, некроз тканей могут вызвать только тахизоиты. Поэтому, когда некоторые авторы пишут, что некроз вызван брадизоитами, то это свидетельствует лишь о том, что пишущий доктор не понимает, чем отличается тахизоит от брадизоита.
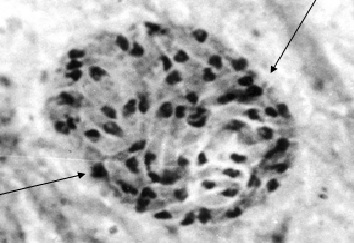
Фото 35 . Мазок - отпечаток головного мозга белой мыши экспериментально заражённой слабовирулентным штаммом токсоплазм «LEI». Показана группа внеклеточных брадизоитов при формировании цисты. Видны брадизоиты имеющие разную морфологическую структуру и находящиеся на разных стадиях деления. Оболочка вокруг колонии ещё отсутствует. Окраска железным гематоксилином.
В отличие от тахизоитов, по мере размножения и формирования колонии, в цитоплазме некоторых видов токсоплазмид, происходит постепенное накопление амилипектина, что меняет не только их морфологию, но и биологическое предназначение. Например, у Besnoitia besnoiti расположенных на периферии колонии, накопленный в цитоплазме брадизоитов амилопектин, идёт на построение внутренней оболочки цистной стенки (Фото 36).
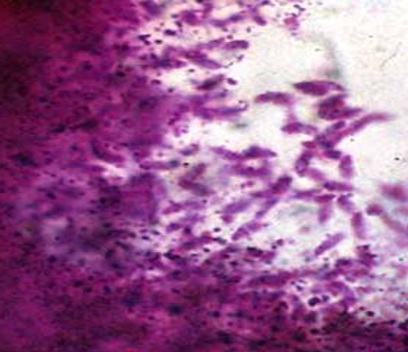
Фото 36. Гистологический срез кожи бычка, подострое течение безноитиоза. Показан участок начала формирования внутренней мембраны цистной оболочки Besnoitia besnoiti. Хорошо виден распад брадизоитов на периферии колонии и высвобождение при этом гранул амилопектина. Окраска по Мак – Манусу.
Не исключено, что и у токсоплазм в образовании оболочки цисты принимает участие, как гидролизированная ткать белого вещества головного мозга, так и амилопектин накопленный брадизоитами. На это указывает положительная реакция оболочки цисты по Мак – Манусу в начальных фазах её образования (Фото 37).
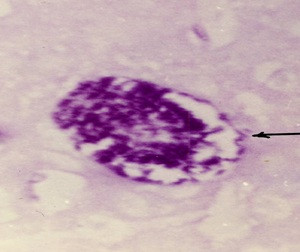
Фото 38. Гистосрез головного мозга белой мыши, на 15 сутки после внутримышечного заражения слабовирулентным штаммом LEI. Вокруг колонии токсоплазм хорошо видна тоненькая, Шик - положительная оболочка. Окраска по Мак – Манусу.
В более зрелых цистах оболочка цист токсоплазм меняет свой химический состав и при окраске по Мак - Манусу уже не даёт позитивную реакцию на амилопектин, а окрашивается базофильно (Фото 39).
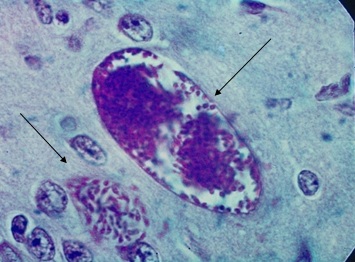
Фото 39. Гистологический срез коры головного мозга белой мыши на 20 сутоки после внутримышечного заражения слабовирулентным штаммом LEI. Показана хорошо сформировавшаяся внеклеточная циста, с чётко видимой тонкой базофильной оболочкой. Рядом, в белом веществе, видна небольшая внеклеточная колония токсоплазм. Цитоплазма брадизоитов даёт интенсивную реакцию на амилопектин. Окр. По Мак - Манусу.
Окраска гистологических препаратов по Маллори, с применением фосфорновольфрамового гематоксилина показывает, что при образовании оболочки цист, принимает участие и гидролизированная гомогенная (слизистая) масса. По методу Маллори оболочка цист даёт реакцию характерную для гидролизированной ткани и окрашивается в тёмно - синий цвет. При этом необходимо почеркнуть, что при окраске гистосрезов по этой методике, в синий цвет окрашивается и колаген. Однако для появления коллагена вокруг колонии токсоплазм нет никаких оснований, поскольку вокруг формирующихся цист никакой клеточной реакции не отмечается. (Коллаген, как известно, образуется фиброцитами, а их вокруг колоний токсоплазм нет, прим. авторов). Интересно отметить, что кариоплазма брадизоитов окрашивалась по Маллори в светло – розовый, нуклеолы в ярко оранжево – красный цвет, а цитоплазма - в светло - синий (Фото 40).
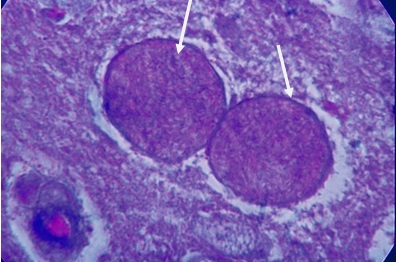
Фото 40. Гистологический срез головного мозга белой мыши на 20 сутки после внутримышечного заражения слабовирулентным штаммом LEI. Показана реакция оболочек цист токсоплазм и самих цистозоитов при окраске среза по методу Маллори с фосфорновольфрамовым гематоксилином. Хорошо видна тёмно – синяя окраска оболочек цист, светло – синяя цитоплазма и ярко -оранжевая окраска нуклеол цистозоитов.
6. Цистозоиты - это медленно размножающие паразиты, находящие внутри цисты. Термин, цистозоит указывает на место нахождения паразита и может быть применим, когда необходимо подчеркнуть, что брадизоит находится в хорошо сформированной цисте. Цистозоиты и брадизоиты отличаются от тахизоитов или трофозоитов, как темпами размножения, так и содержанием в их цитоплазме амилопектина. Если в цитоплазме трофозоитов токсоплазм преобладает РНК, то в брадизоитах амилопектин (Фото 41 - 42). В последние годы установлено, что они отличаются и по антигенной структуре.
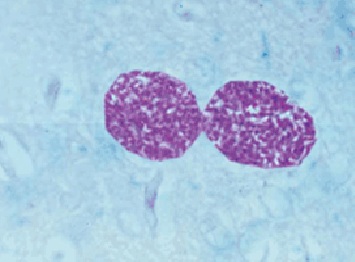
Фото 41. Гистологический срез головного мозга белой мыши заражённой токсоплазмами слабовирулентного штамма токсоплазм. Показаны две хорошо сформированные цисты, оболочка цист окрашена базофильно, тогда как цистозоиты дают резко положительную реакцию на амилопектин. Окраска по Мак - Манусу.
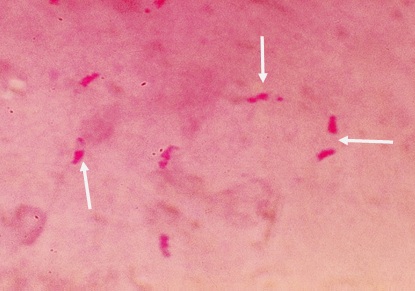
Фото 42. Мазок -отпечаток головного мозга белой мыши заражённой слабовирулентным штаммом токсоплазм. Показано неравномерное распределение амилопектина в цитоплазме цистозоитов. У одних цистозоитов амилопектин расположен большей частью в задней части тела, у других он виден как в задней, так и в передней части паразитарной клетки. Окраска по Мак – Манусу.
8. Оболочка цист токсоплазмид - это сложно организованная внеклеточная структура колонии паразитов, обеспечивающая защиту цистозоитов от агрессивного воздействия организма и способствующая питатанию многотычячной колонии паразитов. Оболочка цист у токсоплазмид образуется тремя путями:
Первый – это трансформация материнской оболочки паразита в оболочку цисты при эндодиогении, эндополигении и шизогонии; Второй – это формирование оболочки из гидролизированных тканей организма хозяина и амилопектина самих паразитов; Третий - часто наблюдаемый у Besnoitia besnoiti - это трансформация ядерной оболочки(ядерной псевдоцисты) в оболочку цисты.
Особо необходимо подчеркнуть, что грануляционная ткань к оболочкам цист токсоплазмид никакого отношения не имеет. Поэтому оболочку цист токсоплазмид лучше не называть капсулой. Поскольку термин «капсула» в организме животных подразумевает образование соединительнотканного происхождения. Например, капсула вокруг небольшого инородного тела или соединительнотканная капсула вокруг личинок гельминтов (халикозы), или соединительнотканная капсула вокруг первичного туберкулёзного очага и т.д.
9. Псевдоциста - это одна из фаз внутриклеточного развития токсоплазмид, чаще в клетках РЭС. Псевдоциста – это не просто клетка инвазированная паразитами. Это внутрицитоплазматическая или внутриядерная колония паразитов, образовавшаяся из одной материнской клетки путём шизогонии, эндодиогении, или полиэндогении. Псевдоциста образуется только в одной паразитофорной вакуоле, в образовании которой важное место занимает оболочка цитоплазмы или ядра инвазированной клетки (Фото 43 – 44). Это последняя стадия бесполого, внутриклеточного размножения токсоплазмид наблюдаемая в подострую и хроническую фазу болезни животных. За этой стадией следует фаза внеклеточного размножения и развития токсоплазмид завершающаяся образованием тканевых цист размножения. Как правило, псевдоцисты не превращаются в цисты. По мере накоплания паразитов в цитоплазме, оболочка инвазированной клетки разрушается, и паразиты покидают погибшую клетку. В псевдоцистах паразиты накапливают питательные вещества для дальнейшего внеклеточного размножения в головном мозге, мышцах и других органах, в которых они формируют тканевые цисты размножения.
В отдельных же случаях, например, у Besnoitia besnoiti, ядерная псевдоциста превращается в тканевую цисту. При этом происходит полная атрофия кариоплазмы и цитоплазмы инвазированной клетки. Оболочка же ядра трансформируется в оболочку цисты. Такая оболочка имеет уже другую структуру и функциональное назначение, а именно она обеспечивает, с одной стороны защиту колонии паразитов от патогенного воздействия организма хозяина, с другой – способствует процессам ассимиляции и диссимиляции тысячной колонии паразитов находящихся внутри цисты.
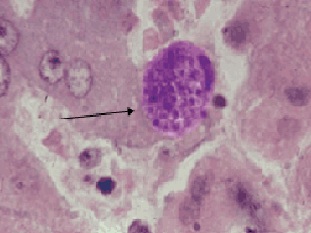
Фото 43. Гистологический срез печени щенка, экспериментально заражённого вирулентным штаммом токсоплазм CDN. Показана типичная псевдоциста в клетке Купфера. Окр. Железным гематоксилином и эозином.
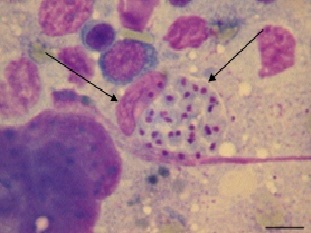
Фото 44. Фотография взята из работы Alexandre Paulino Loretti, и Severo Sales Barrosb «Hemorrhagic disease in dogs infected with an unclassified intraendothelial piroplasm in southern Brazil», опубликованной в 2005 году в Бразилии.
Показана типичная псевдоциста в эндотелиальной клетке почки (клетке РЭС) при экспериментальном пироплазмозе собаки.
(Bone marrow, dog, experimental case. Smear. Many zoites are found in the cytoplasm of an endothelial cell. Panoptic. Bar = 4.5 μm)
10. Инвазированная клетка – При остром течении токсопдазмоза, в гистологических срезах, мазках - отпечатках из органов, в мазках крови, в мезотелиальных клетках брюшины и плевры можно обнаружить множество инвазированных клеток. Нередко клетки одновременно инвазируются несколькими трофозоитами, которые дают начало как бинарному, так и множественному делению (шизогонии). Результатом такой инвазии является гибель клетки и образование микронекрозов с многочисленными паразитами вокруг некротизированной ткани. Клетка инвазированная токсоплазмами и особенно множеством токсоплазм, никогда не превращается в псевдоцисту, она разрушается, а паразиты выходят в межклеточное пространство и дают начало новому поколению паразитов. Нередко, вместе с распадом инвазированной клетки погибают и сами паразиты. Если инвазии и разрушению подвергаются мезотелиальные клетки то в брюшной экссудат выходят паразиты, находящиеся на разных стадиях развития и размножения (Фото 45 - 46).
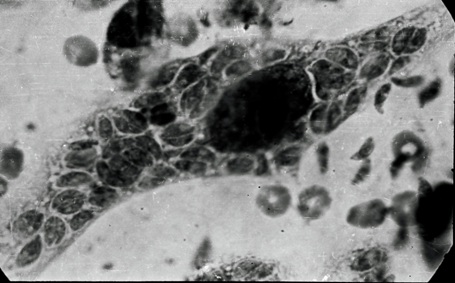
Фото 45. Пластинчатый препарат брыжейки белой мыши. Показана мезотелиальная клетка при одновременной множественной инвазии. Токсоплазмы находятся на разных стадиях бинарного и множественного деления. Вокруг каждой группы паразитов имеется своя паразитофорная вакуоль. Окраска по Романовскому - Гимза.
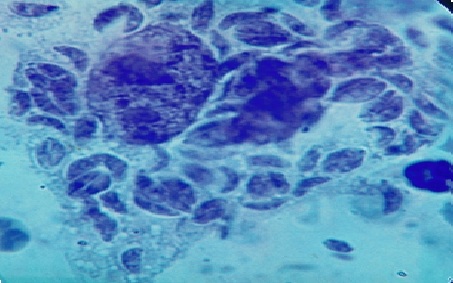
Фото 46. Пластинчатый препарат брыжейки белой мыши павшей на 6 сутки после заражения. Показана мезотелиальная клетка, одновременно инвазированная множеством токсоплазм. Цитоплазма инвазированной клетки вакуолизирована, местами оболочка клетки разрушена, а паразиты свободно покидают клетку. Окр.железным гематоксилином и эозином, с подкраской метиленовым синим.
10. Мерозоит - это стадия жизненного цикла паразитов образовавшихся в результате шизогонии.
11. Зигоциста – это неспорулированная зигота. Зигоцисты, выделяются во внешнею среду, вместе с калом, после половой фазы размножения токсоплазмид в эпителии тонкого отдела кишечника, представителей семейства псовых и кошачьих.
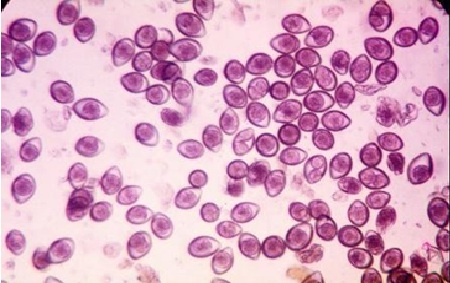
Фото 47. Зигоцисты (I.felis), в кале пятимесячного котёнка через 4 часа после выделения их во внешнюю среду. Большая часть зигоцист не спорулирована. Естественное заражение.
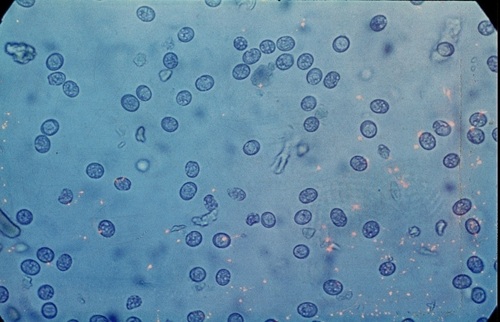
Фото 48. Показано больше количество зигоцист токсоплазм в кале экспериментально заражённого котёнка через 3 минуты после выделения их из кишечника. Такие зигоцисты не опасны для других животных и человека.
Зигоцистами заразится не может ни человек, ни животные, они не инвазионные (не заразные) образования. В них ещё нет спорозоитов, которве должны образоваться при споруляции, во внешней среде.
Термин зигоциста для токсоплазм ввёл академик И.Г.Галузо, чтобы подчеркнуть разницу между токсоплазмами и кокцидиями. Однако этот термин правомерно может применяться и для других паразитов образующих изоспоры. Зигоцисты во внешней среде, при хорошем доступе кислорода спорулируют и становятся инвазионными (заразными).
|
|
13.Изоспора – это спорулированная зигоциста. В результате споруляции в изоспоре образуется две споры с четырмя спорозоитами в каждой. Изоспоры образуются во внешней среде, при доступе кислорода через 15 – 20 минут после попадания их во внешнию среду! В некоторых работах приводятся данные, что изоспоры образуются через 5 – 6 часов - это может связано с плохой аэорацией зигоцист. В нормальных домашних и природных условиях споруляция начинается уже через 15 – 20 минут. Учитывая такую скорость споруляции зигоцист токсоплазм, для собственной безопасности, кал кошек необходимо убирать как можно быстрее.
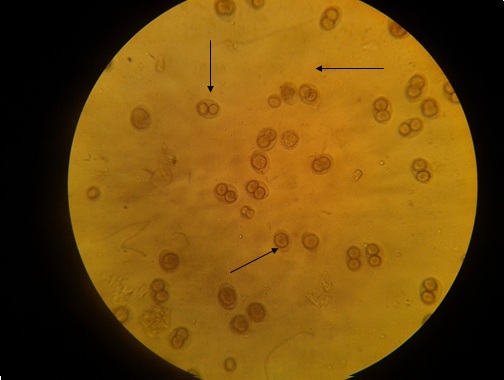
Фото 49. Показаны спорулированные изоспоры ((I.felis) и Т.gondii)
выделенные из кала 2 –х месячного котёнка при смешанной инвазии.
11.Амилопектин – Один из основных полисахаридов крахмала, состоящий из разветвлённых цепочек молекул глюклзы. Он служит главным питательным и структурным материалом для токсоплазмид, особенно в фазу образования псевдоцист и тканевых цист размножения. Роль амилопектина в разных стадиях развития токсоплазмид точно не установлена. Если у токсоплазм и безноитий, по мере их внутриклеточного размножения, амилопектин накапливается в цитоплазме паразитов и максимум его отмечается в брадизоитах, то у саркоспоридий формирующих цисту или находящихся в цистах амилопектин отсутствует.
ПРИМЕЧАНИЕ: Все фотографии, где не указано авторство, принадлежат лично А.М. Бугаеву и находятся в архиве Центра охраны здоровья животных г. Киева. Под фотографиями указаны увеличения, которые сделаны при съёмке объекта, однако многие размеры фотографий были увеличены или уменьшены с помощью компьютерной техники и могут не соответсвовать истенным увеличениям.
Телефон для контакта - (044) – 426 – 47 – 76 (78).
Email: bugaev@ukr.net
- © 2013-2015 www.vetbugaev.kiev.ua
- Все права защищены.
- Использование любых материалов, размещённых на сайте, разрешается при условии ссылки на www.vetbugaev.kiev.ua
- При копировании материалов со страницы www.vetbugaev.kiev.ua для интернет-изданий – обязательна прямая открытая для поисковых систем гиперссылка. Ссылка должна быть размещена в независимости от полного либо частичного использования материалов. Гиперссылка (для интернет- изданий) – должна быть размещена в подзаголовке или в первом абзаце материала.
Обновлено (24.01.2019 09:31)